








Takaisin Ajatusvarikolle
-
Back to the Thought Deposit
Dinoglyyfit
-
Dinoglyphs











SYMBIONTTI-lehti (Symbioosi ry)
Lajien loppu – dogmien synty?
Osa II: Ajan lyhyt hysteria
The Extinction of Species & Origin of Dogma - The Short Hystery of Time
"Scarlett O'Hara ei ollut kaunis, mutta sitä harvoin tulivat huomanneeksi ne, jotka joutuivat hänen viehätysvoimansa lumoihin..."(1) Myös tieteiden keisari on kohtalokas paradigma. Humanismin yhtenäisteoriaa evoluutiota vaivaa kehäpäättelyn ongelma, jonka oivaltaa syvällisesti Mika Häkkisen oranssien/sinisten aurinkolasien suoman metaforan kautta. Koska hysteria ruokkii itse itseään, voidaan kysyä: ollaanko lajien muuntelukapasiteetin suhteessa ajanjaksoihin menetetty suhteellisuudentaju?
Keskimäärin yhden lajin päivässä
arvellaan kuolleen sukupuuttoon vuodesta 1758, josta lähtien eläinlajien
tilastoja on pidetty. Vuoden 1998 IUCN Worldwatch-raportin mukaan suurista
eläimistä parhaiten tunnetaan linnut, joiden lajimäärä
on 10 000 tietämillä ja joista uhanalaisia on yli tuhat. Matelijoita
lienee niin ikään 10 000 paikkeilla, ja nisäkkäitä
on noin 4400, joista neljänneksen olemassaolo on vaarassa. Matelijoista
joka viides, sammakkolajeista joka neljäs ja kaloista joka kolmas
on tänään uhanalainen.
Mitä kasvillisuuteen tulee, niin viimeisen sadan vuoden aikana puolet maapallon kosteikoista on tuhoutunut ja puolet metsistä kaadettu. Maapallon maapinta-alasta (jää ja kallio poislukien) häiriytymätöntä on 27 %. USA:ssa luonnonmetsistä on jäljellä 15 %, Kanadassa 40 %, Euroopassa ja Kiinassa 1 %. Alkaneen vuosisadan aikana ennustetaan Telluksen eliölajeista puolen kuolevan sukupuuttoon. Nopeuden uskotaan tätä nykyä olevan tuhatkertainen fossiiliaineistosta pääteltyyn taustaan nähden (2). Tätä vauhtia joissakin tuhansissa vuosissa päästäisiin bakteerien valtakauden tilastopoikkeamista kokolailla kokonaan.
Teknokratian käppyrät ovat kyllä tempaisseet viimeisten parin sadan vuoden aikana eksponentiaalisiksi, mutta jospa luteita rassaa myös kertaluokkien ero geologisen ja biologisen ajanmäärityksen välillä? Entropian saldo pääomittuu WWF:n rahastoa varmemmin, koska termodynamiikan toinen pääsääntö on loppujen lopuksi ns. luonnonlaki.
Pohjois-Amerikassa eli vielä alle 10 000 vuotta sitten parhaimmillaan seitsenmetrinen jättiläislaiskiainen ja Etelä-Amerikan intiaanit päällystivät kolmimetristen vyötiäisten panssareilla hautojaan. Moa oli lentokyvytön, strutsin kaltainen lintu joka kasvoi jopa kolmemetriseksi ja joka metsästettiin sukupuuttoon vasta noin 300 vuotta sitten. Jopa 450-kiloinen Marco "miljoona" Polon nimeämä Madagasgarin norsulintukin eli vielä 1600-luvulla. Monet Pohjois-Amerikan intiaaniheimot tunsivat parhaimmillaan kolmanneksen nykynorsuja suuremmat mammutit ja vuonna 1873 raportoitiin vielä silminnäkijöiden väitteitä Siperian mammuttilaumoista (3). Venäjän tasangoilla ihmiset jopa asuivat mammutin luista tehdyissä majoissa. Kolmeen kivikautiseen taloon Ukrainassa käytettiin 1700 luuta jotka olivat peräisin vähintään sadasta mammutista. Ensimmäisenä Euraasian ympäri purjehtinut nationalistimme AE Nordenskiöld kertoi pohjoiskalotin asukkaiden mammutinjäämistöistä valmistetuista käyttö- ja taide-esineistä. Hän arveli alueella liikkuneen kauppatavarana yli 20 000 mammuttiyksilön jäämistöä (4).
Antiikin semi-eläintieteellisiin klassikoihin tutustuvalle
voi tulla yllätyksenä, miten paljon niiden kuvaamista eliöistä
on kuollut sukupuuttoon historiallisella ajalla. - Siitäkin huolimatta,
että luokittelisi oheisten viitteiden vanhojen partojen "lentävien
käärmeiden", ukkoslintujen, lohikäärmeiden ja muiden
hirmuisten liskojen kerettiläisen kategorian satujen taksonomiaan
(5). Kotopuolessahan nämä myytit kulkivat vaakalinnun, tursaan
tai ikiturson nimillä muinaiskalevalaisessa aineistossa. Nämä
nuotiojutut ovat sikäli erikoisia, että esiintyvät paisutteluista
huolimatta samankaltaisina Japanista aina Keski-Afrikkaan, Lähi-Itään,
Eurooppaan ja Amerikkoihin saakka. On harmillista, ettei Lemminkäisen
Hiiden hirven hiihtoa tai –nelimetrisiä sarvia kantaneen saksanhirven
fossiiliaineistoon sopivammin – Nibelungin hirveä ole dokumentoitu
riittävällä tarkkuudella.
Kellojen kalibroinnista
Kestävimmänkin kehityksen periaatteen tuloksena uhkaa virhekatastrofi ("mutational meltdown"), jos sille annetaan äänekkään kysynnän vaatimat ajanjaksot. Lukuisista korjausmekanismeistä huolimatta meioosilla on jo pohjalla peitettävänään kummankin ihmisgameetin vähintään 10-20 mutaatiota. Mutaatiokellohypoteesissa on suuria epätarkkuuksia, kuten Simon Conway Morrisin vuosituhannen ensimmäisen, satalitraisen Cell-lehden, ensimmäisen jutun introssa todetaan: "Evolution has a temporal framework, but molecular ’clocks’ now plot a history of life seriously at odds with the fossil record. Which is correct?" (6)
Y-kromosomin paternaalinen ja monikopioisen mito-DNA:n (lähes) täysin maternaalinen periytyminen luo mahdollisuuden mutaationopeuden tarkemmalle määrittämiselle. Solun jakautuessa sen sadoista mitokondriosta vain kymmenisen kappaletta periytyy pullonkaulana eteenpäin. Mito-kromosomin erikoisuuksiin kuuluu vielä intronien ja histonien poissaolo. Mito-DNA-tutkimuksessa on eletty jo 20 vuotta "postgenomista aikakautta", mutta näihin vuosiin asti nopeus on laskettu jakamalla sekvenssieron määrä eliöiden geologisen iän erotuksella, eikä kukaan ole oikein vaivautunut askartelemaan todellisen muutosnopeuden kanssa.
Ensimmäiset mtDNA-tutkimukset, jotka käyttivät oletuksenaan erilaisten saarten arvioituja asuttamisajankohtia, tiputtivat ihmiskunnan ikää noin 200000 vuoteen (7). Mt-DNA:n suojausmenetelmien tiedettiin jo etukäteen olevan tuman DNA:han verrattuna lähes olemattomia, mutta siitä huolimatta ihmisen pullonkaulapopulaation "Eevan" alustava kellotus lähelle historiallista aikaa tuli out of the blue (8). Parsons ym. jättivät olettamukset minimiin ja tutkivat 134 toisilleen kaukaista mammalinjaa yhteensä 327 sukupolven vaihdoksessa. Kyseisessä artikkelissa mainitaan tosiaankin nelinumeroinen luku ilman eksponentteja ihmiskunnan MRCA (most recent common ancestor) –arvona, joskin tekijät kiirehtivätkin heti tämän sanottuaan varoittelemaan tuloksesta vedettävistä hätiköidyistä johtopäätöksistä. Mt-DNA:n 614 nukleotidin säätelyalueella tapahtuu tämän mukaan 1/33 muutosta generaatiossa eli 2,5 muutosta per nukleotidi per jokainen hihasta vedetty vuosimiljoonaa.
Edellistäkin paremmaksi pistivät Denver ym. syyskuun lopulla, kun Caenorhabditis eleganssin keskimäärin 214 sukupolven (a’ noin 4 vrk) saaduksi vastaavaksi emästen substituutioarvoksi saatiin 8,9 per nukleotidi per ekstrapolaatio miljoonaan vuoteen! Aika eleganttia siihen nähden, että ero vakiintuneeseen käsitykseen on satakertainen nopeampaan suuntaan (9). Ryhmä sekvensoi 74 fletkumato-linjaa pisimmän alaan liittyvän kasvatuksen jälkeen koko genomin 10 kb matkalta – ja kaikki tämä mahdollisimman seesteisissä kasvuolosuhteissa. (Jolloin mutaatioita toisaalta tapahtuu vähemmän ja toisaalta ne säilyvät populaatiossa pienemmässä selektiossa kauemmin. Ihmisenkin mt-DNA:n pituus on vain 16,6 kb.)
Vuoden lopulla kohun kanssa julkaistu koko mitogenomin kattanut ihmisheimojen kartoitus laski Afrikan exoduksen ajankohtaa 52000 vuoteen 28000 vuoden virhemarginaalilla (10). On kuitenkin merkillepantavaa, että tässä raportissa huomio keskitettiin tasaisella nopeudella muuttuvaan mito-sekvenssiin. Työssä siis jätettiin paitsioon nimenomaan em. eniten varioiva D-luuppi, joka kattaa 7 % omasta mitogenomistamme ja antoi siis aiemmin kertaluokkaa pienemmän iän. Nopeutta ei lisäksi käsittääkseni kalibroitu äitien ja tytärten eroilla, vaan sillä suht puhtaalla oletuksella, että ihminen ja simpanssi erosivat toisistaan 5 Mrv sitten - seikka joka siirtää työn semisarjaan empiirisyydessään. Tässä ruotsalaisten tekemässä raportissa kiinnostavaa on mielestäni pullonkaulapopulaatioiden lukumäärän asettuminen kolmeen. Tutkimus tukee lisäksi vanhaa "Out-of-Africa-Vähä-Aasian-vuoristoseudun-kautta"-teoriaa. Työn pohjalta voisikin leikillään esittää uutta MRCR-käsitettä, jossa viimeinen kirjain tulee sanasta route.
Jo ennen näitä töitä Svante Pääbo ilmaisi huolensa seuraavin sanoin: "…the high mutation rate represents an impressive discrepancy with the received wisdom among evolutionists" (11). Diskrepanssia on kuitenkin puolusteltu sillä, että mitokondrion muutokset ovat biosfäärin kellon minuutti- tai (D-luupin tapauksessa) sekuntiviisari. Viisareilla on kuitenkin se piirre, että ne saavat uuden tilaisuuden aina 60 yksikön välein. Satunnaisella mutaatioihinsa hirttyvällä organismilla tätä luontaisetua ei ole, joten tämä vertaus on vain silmänkääntötemppu. Paitsi että mitokondrioiden mutatoituminen voi tappaa sukupuuttoon, se saattaa ehtiä tappamaan ihmisyksilönkin perinnöllisten sairauksien ja vanhenemisen kautta. Sirkulaarisesta mitogenomista voi kuulemma irrota kokonaisia paloja tai deletoitua geenejäkin. Pääboon oma neanderthalin ihmisen mt-sekvensointikin näyttää tulleen juhlituksi liian aikaisin siinä uudessa valossa, että tätä nuoremmat nykyihmisjäänteet Australiasta ovat sitäkin kauempana nykypopulaatiosta. (Ja viittaisivat itse asiassa ihmiskunnan syntyneenkin Australiassa…12)
Pappa-puolella laajojen otosten Y-kromosomin kartoitukset todistavat eksponentiaalisesta väestönkasvusta, joka alkoi alle 30 000 vuotta sitten (13). Viime aikoina ollaan päästy vetämään johtopäätöksiä myös kokonaisvaltaisissa genomianalyyseissä viidenkymmenen eubakteerin ja arkin sekvensointiprojektien purkittuessa. Pöpöt antoivat karun tuloksen, jonka mukaan graduaalista ja yleistä fylogeneettistä signaalia ei pääosalle proteiiniperheistä ole olemassakaan. Tuomas seisoo niin kuin tippaleipä. Elämän puu paljastui "elämän puskaksi" (14, 15). Runollisemmin ilmaistuna: elämän puu on kaatunut. "Horisontaalinen" tai "lateraalinen geenisiirto" (LGT) osoittautui taikasanaksi tai matoksi, jonka alle kiusalliset konvergenssit lakaistiin. "Elämän puun" ulkonäkö riippuu siitä, minkä geenin perusteella sen rakennamme (16, 17).
Mitä "korkeampiin" aitotumaisiin eliöihin ja
rappeutumiseen tulee, niin ihmisgenomihankkeen yhteydessä toistettuja
sanoja ovat mm. hautausmaa, transposoni, pseudo, parasiitti, virus ja moska.
Massiivisen hankkeen valmistuessa muutenkin raadollisissa merkeissä,
herää kunnallispolitiikan syrjästäkatsojan mielessä
pakostakin tuo alunperin HC Andersenin muovailema kysymys: onko
tieteiden keisari ollenkaan falsifikoitavissa? Tilanne muistuttaa surullisen
paljon erästä toista rahahanketta, kun Neil Armstrong Kotkan
laskeutumista seuranneessa hässäkässä lausui jotenkin
kevyet sanat suuresta askeleesta ihmiskunnalle - tuuman verran pölyyn
uponneen askeleen johtopäätökset olisivat voineet olla painavammatkin
(40).
Toisinaanajattelijan hakusanoja
Replikaation virheiden on perinteisesti katsottu olevan mutaatioiden pääasiallinen lähde. Luonnossa esiintyy kuitenkin vain harvoin eksponentiaalista kasvua. (Esimerkiksi E. coli jakaantuu suolistossamme pari kertaa vuorokaudessa eikä suinkaan tunnissa.) Mitä spontaaneihin mutaatioihin tulee, niin on viitteitä, että mutaatioissa on spesifisyyttä, joka suistaa tilastolliset arviot sivuun. Aiheen hakusanoja ovat sekventiaaliset, toistaan systemaattisesti seuraavat mutaatiot, hot-spotit, adaptiiviset mutaatiot, fenotyyppinen plastisiteetti – ja kuriositeettina prionit signaaliantureina. Ympäristön vaikutus niin feno- kuin genotyyppiinkin on jälleen kiinnostava kysymys, vaikka vain harva kannattaakaan yleisnero Lamarckin kaivamista ylös haudastaan.
Virikkeiden päällekytkemät latentit kapasiteetit
ja lyhyemmät aikajaksot pienentävät sattumanvaraisen evoluution
mahdollisuuksia informaation moottorina ja vahvistavat käsitystä
sisäänrakennetuista muuntelukoneistoista. Sokea kelloseppä
on joko käyttänyt luuppia, tai sitten luonto kiittää
luonnonystävää.
Kambrin informaatioräjähdys
Todellinen alkuräjähdys oli biologinen. Kambrin informaatio-räjähdyksessä fossiilirekordiin ilmestyy paria poikkeusta lukuunottamatta noin 550 Mrv (miljoonaa radioaktiivista vuotta) sitten (3500 Mrv yksisoluisen ajan jälkeen) mahdollisesti jopa 5-8 Mrv:ssa kaikki noin 33-39 nykyisin tuntemaamme eläinten pääjaksoa. Itse asiassa alussa oli mukana lisäksi 20-70 sukupuuttoon kuollutta pääjaksoa - näköjään homeodomeineen kaikkineen – joskin jokerien lukumäärästä kiistellään (18). Prekambrin (rifean, vendian) monisoluisten eläinten olemassaolo on jossakin määrin kiistanalainen, mutta ainakin joitakin sienistä on viitteitä.
Asian varmemmaksi vakuudeksi lainattakoon tässä muutamaa asianharrastajaa: "The ’Cambrian explosion’ is a real evolutionary event, but its origins are obscure. At least 20 hypotheses have been proposed, and although arguments linking diversification to oxygen levels, predation, faunal provinciality and ocean chemistry all attract support, it is he case that ’The emergence of Metazoa remains the salient mystery in the history of life’" (19). "Phyla appear abruptly in the fossil record without intermediates to link them to their putative ancestors. This pattern presumably reflects derivation of most or all phyla from small, soft-bodied ancestors that had virtually no potential for fossilization. However, most classes and orders of durably skeletonized marine animals also appear abruptly, without obvious linkage to their durably skeletonized antecedents…(20)". "…the extreme speed of anatomical change and adaptive radiation during this brief time period requires explanations that go beyond those proposed for the evolution of species within the modern biota" (21).
Kambista on naarattu mm. pari omaan selkäjänteisten pääjaksoomme kuuluvaa kalaa loisineen kaikkineen (22). Koska räjähdyksen lajit ovat lisäksi maantieteelliseltä korkeudeltaan meren eliöstöä, on koko monisoluisen elämän kirjo kalkkiviivoilla avoin tapaus. Villaa puolet, loput viskoosia.
"Ylläpito-domeenit" ovat transkriptiofaktoreissa ym. esiintyviä standardeja DNA:ta sitovia laskoksia, tai "geenikatkaisimia", jotka ohjailevat elinten kehitystä kuin metalliosat sateenvarjoa. Harhaanjohtavasti "makromutaatioiden" aiheuttajana markkinoituja homeodomeeneja on löytynyt kaikista tähän mennessä tutkituista monisoluisista organismeista ja lähinnä niiden lukumäärä korreloi eliöiden kompleksisuuteen.
Anterior-psterior -askelia kontrolloivien Hox-geenien manipulaatiolla saadaan esim. raaja ilmentymään eri paikkaan kehoa. Muita geenejä ovat mm. fringe (raajat), tinman (sydän) ja Pax6 (silmä). Homeodomeenien havainto vähentää sattumanvaraisen informaatioräjähdyksen todennäköisyyttä entisestään, koska näillä domeeneilla on merkitystä vain, kun ne tarttuvat kunkin pääjakson ominaisuuksien kirjon aikaansaaviin geeneihin sateenvarjon metalliripojen tavoin.Pelitaktiikkaa luulisi olevan vaikea suunnitella ennen kuin kortit on jaettu tai komppaniaa organisoida ennen aselajin valintaa.
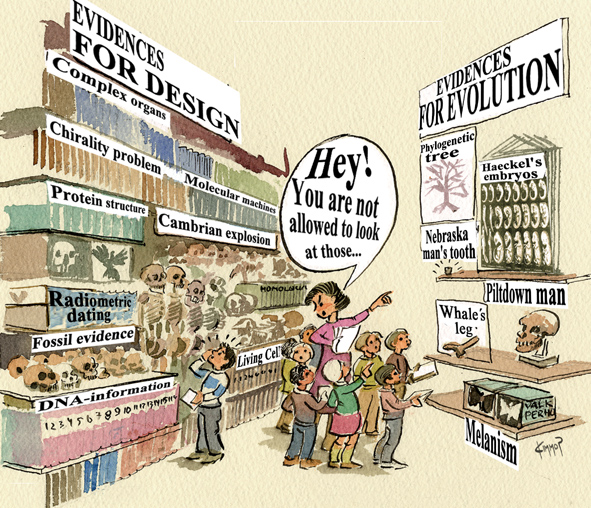
On mielestäni oireellista, että lukion oppikirjat jättävät "Kambrin räjähdyksen" systemaattisesti joko vähälle huomiolle tai sitten kokonaan käsittelemättä. Yksi asia on, jos yleisölle ei esitellä kuin yhtä tulkintamallia, toinen jos havainnot (puuro) ja tulkintamalli (velli) sekoitetaan, ja kolmas jos kiusallisesta havaintomateriaalista ei edes kerrota. Tieteellisen teorian pahin vihollinen ei ole vastustus, vaan se, mitä immunologit kutsuvat nimellä "clonal anergy" tai minkä englantilaiset sanalla "ignore" ilmaisevat.
"Paleontologien likaisen pikku ammattisalaisuuden" eli ylimenovaiheiden poissaolon möläyttänyt Gould pyörittää samoja parametrejä Dawkinsin "epätodennäköisyyden vuorta" punktuuratessaan. Pullonkaulasta syntynyt populaatio ei välttämättä kuole sukupuuttoon, mutta "siirretyn tasapainotilan" yhtälöillä on se erikoisuus, että niiden päätodiste on (fossiili)todisteiden puute.
Tässä turhan useasti enemmän lämpöä kuin valoa tuottavassa keskustelussa x-akselin muuttujana on gradualistisen kehityksen osuus ja y-akselilla juuri tämä punktualistinen "toivottujen hirviöiden" osuus. Tasoon painetusta kirjallisuudesta puuttuvana z-akselin parametrina on katastrofismi ja sen maantieteellinen laajuus. Aika on neljäs ulottuvuus, mutta sitä näköjään tulee kuin siimaa jos kysyntää ilmenee. Ns. "megasukkession" haastajateoriassa geologiset kerrostumat ovat vähemmän kello ja enemmän ekologinen postilokero (23).
Varhaiskambrikerrostumista löytyy fosfaattipitoisia small-shelly-fossiileja fosfaattipitoisten sedimenttien kanssa, ja rakennusmateriaalin hävitessä häviävät myös sille ominaiset eliötyypit. Fosfaatin jälkeen ensisijainen rakennusmateriaali puolestaan on karbonaatti. Valistuksen ajalta lähtien taakseen poistetun katastrofismin voisi tuoda takaisin selittämään sitä, etteivät ainoastaan eläinhahmot ilmesty esiin yhtäkkiä ja häviä jälleen yhtä nopeasti, vaan sama tapahtuu kokonaisille monimuotoisille ekosysteemeille.
Jos aika on neljäs ulottuvuus, niin räjähdyksemme ajankohta määritettiin alunperin sedimentaatiokerrosten paksuuden perusteella ennen nk. absoluuttisten radiokellojen keksimistä. Kun tämä toinenkin sääty sai kellon, antoi se miellyttävästi saman ajan. Radiokellon ongelma on kuitenkin oletukset hajoamisketjujen aineiden määristä lähtöhetkellä vulkaanisen tapahtuman jälkeen. Havaintojen kertyessä geologisten kerrostumien todellinen paksuus on kuitenkin melko epämääräinen. Oppikirjojen reilut sata kilometriä on saatu summattua palapelityyliin. Kelloa on noloa rukata, joten geologit luottavatkin fyysikoiden "absoluuttiseen" kelloon. Sopimukselta haiskahtavaa kellojen naimakauppaa on vaikea tarkistaa, koska geologinen kokonaispaksuus elää omaa elämäänsä. Kuten hiljaisella tulella sammakko kellahtaa vesikattilassa selälleen, tulee vaihtolämpöisestä teoriastakin varsin tasalämpöinen suhteellisuudentajun saavutettua sadan asteen etapin.
Tårta på tårta. NDT (neo darwinian theory)
on pahimmillaan tautologista kehäpäättelyä, jossa kelpoisin
tarkoittaa sikiävintä, ja paleontologit johtofossiileineen sekä
fyysikot radiokelloineen tarraavat toisiinsa kuin veljekset DuPond ja DuPont
Tintin kuuraketissa maan vetovoiman sammuessa. Kolmannen säädyn,
molekyylibiologien, saatua kellon, voidaan siis todeta: OK – "All Correct":
kellot ovat ristiriidassa keskenään (41).
Muinaismolekyylit
PCR-menetelmän kehittämisen jälkeen DNA:ta yritettiin pilan päiten monistaa fossiileistakin. Kun tämä sitten onnistuikin, oli ihmetys suuri (24). Asiaa hämmensi Crichton ja Spielberg Jurassic Park –jutuillaan, mutta uuden dinobuumin takana ei siis ole pelkkää fiktiota. Tämänhetkisen mielipiteen mukaan työt on kumottu kontaminaatioriskiin sekä vaikeaan toistettavuuteen vedoten. Hassua on vain se, että kymmenisen kappaletta alan jutuista ehdittiin julkaista parhaissa tiedelehdissä 90-luvun alkupuolella. Tällöin töitä kutsuttiin useamman ryhmän toistamaksi ja sekvenssianalyysin sanottiin varmistavan näytteiden aitouden ja niistä vedettiin johtopäätöksiä kladistiikan hienosätöön. Meripihkamehiläisen tapauksessa ei natsannut ainoastaan hyönteisen, vaan myös sen enterobakteerin sekvenssit (25). Viikin maanantaiseminaarissa vieraillut Svante Pääbo kertoi DNA:n säilyvän korkeintaan 30-50000 vuotta, varsinaiset DNA:n rakenteen tutkijat sanovat joskus katon olevan 10 000 vuotta. Parhaimmissa töissä ei siis ole huomautettavaa tekniikan, vaan teorian puolella (26).
Vaika PCR-tulokset olisivatkin sammakoita, niin DNA:n eristämistä ylipäänsä on kuvattu tätäkin enemmän. Magnolia herätti kiinnostusta, koska kymmeniä miljoonia vuosia vanhan näytteen vihreä väri katoaa joissakin sekunneissa kun fossiili käännetään vanhasta järvenpohjasta ja savilaatasta auringonvaloon. Mitä toistettavuuteen tulee, niin "muinais-DNA" on huonossa kunnossa. Se esiintyy vain muutaman sadan nukleotidin pätkissä, juosteet ovat "takussa" ja sen emäkset ovat usein hapettuneita tai kokonaan poissa. Mm. tammen, tulppaanin, magnolian ja sypressin fossiililehdistä on onnistuttu eristämään DNA:ta ja sekvenoimaan niistä biosfäärin yleisintä proteiinia, kloroplastista monikopioista rubiscoa (27). Vaikka kaikki todella olisi ollut kontaminaatiotakin, on hälyttävää miten havainnot selitettiin sujuvasti pois tuloksiin vielä luotettaessa. Tällä saralla ylpeys ja ennakkoluulo törmäävät kunnolla.
Liitukauden (65 Mrv) tuolta puolen on löydetty myös mm. dentiiniä, kollageenia ja osteokalsiinia (28-31). Kuitenkin 13000 vuoden ikäiseksi arviodusta mammutinluusta kaikki kollageeni on saattanut jo hävitä (32). Koppakuoriaisen kitiini välkehti 25 Mrv jälkeen ilkikurisesti kaikissa pääväreissä ja varmuuden välttämiseksi asia sinetöitiin massaspektrometrillä ampumalla (33). Dinoluut eivät monesti ole kokonaan kivettyneet. 80 Mrv takaa kalsiumin ja fosforin suhde saattaa olla aivan samanlainen kuin nykyisissä luissa (34). Alaskan rannikolla tutkijat eivät koskeneet lähes kivettymättömiin dinosaurusten luihin, koska olettivat kyseessä olleen biisonien luut (35). Kivettynyttä puuta syntetisoidaan bisnes-mielessä ja sitä muodostuu esim. kuumissa lähteissä joissakin kuukausissa. Eteläafrikan boikotin aikaan taas maassa tehtiin pienessä mittakaavassa biomassasta öljyä parhaimmillaan varttitunnissa, kunhan olosuhteet vain ovat kyllin kovat.
PCR-temput ovat uusi menetelmä, mutta mm. 25 Mrv meripihkasta on kyetty eristämään myös eläviä – vieläpä kriitikoille resistenttejä – bakteereita (36). Tämä on varsin erikoista siihen nähden, että aminohappojen rasemisaationopeuksista eri olosuhteissa on aivan eri kertaluokkien empiiristä dataa. L-aminohapot rasemisoituvat peilikuvikseen puoliintumisnopeuden vaihdellessa 400 ja 50000 vuoden välillä (25 oC), sata kertaa hitaammin nollassa asteessa (37). Vaikka itiöissä olisi aineenvaihduntaa hapettumisen, rasemisaation jne. korjaamiseksi, niin tähän tarvittava energialähde puuttuisi. Samat ryhmät ovat herättäneet meripihkafossiileista henkiin tieteelle tyystin tuntemattomiakin bakteerilajeja (38).
Jos siis 569 miljoonaa vuotta vanhasta ja 250 metriä
syvältä - anteeksi toisinpäin - kaivetun suolakiteen vesitaskusta
löytyy eläviä Bacilluksia (39) ja havaintoa diskutoidaan
piristävänä lisätodisteena evoluutiosta, ei kyse ole
tieteellisestä, vaan uskonnollisesta keskustelusta. Jälkimmäinen,
TV-uutisia myöten popularisoitu viime syksyn löytöhän
valjastettiin aihetodisteeksi aurinkokuntien välisestä "panspermiasta",
vaikka 16S rDNA:n perusteella muinaisbaku oli 99 % samanlainen Bacillus
marismortui -halobakteerin kanssa. ("Kuollutmeri" ei siis olekaan kuollut.
Tätä aiemmin ikäennätystä piti siis hallussaan
25-40 miljoonaa vuotta vanhaksi katsottu bakteeri, joka löydettiin
aikoinaan meriphkan sisään umpioituneen mehiläisen sisuksista.
) SETIllä taitaa olla silmät eri indeksillä teleskooppiin
ja mikroskooppiin katsottaessa, mitä älyn ulkopuolisen rahoittajan
olemassaoloon tulee. Evoluutio vaikuttaa liian syvältä paradigmalta
luonnontieteen pitkän kouran ylettyä. Mikäli poikki tieteen
juuttunut teoria ja tulkinnallinen viitekehys unohdettaisiin, olisi tällaisten
johtopäätösten relevanssi tohu ja bohu - autio
ja tyhjä.
Pauli Ojala
http://www.kp-art.fi/taustaa/index.html
Katso myös Duodecim-artikkeli
hirmuliskon
verisuonista
Kuva 2. Nehän ovat kuin kaksi marjaa!
3,4 x 109 Mrv ikäinen Ramsaysphaera ja nykyinen Candida tropicalis.
Evoluutio-kriittinen analyysi s. 212.
Kuva 3. Nehän ovat kuin kaksi marjaa! Magnetobakteerien mikrofossiileja ja
nykyisiä magnetosomeja. Evoluutio-kriittinen analyysi s. 213.
1/Mitchell M (1978) Tuulen viemää, n. painos, WSOY.
2/Ilkka Hanski 29.1.2001 Viikin maanantaiseminaarissa.
3/Johnson, LH Men and Elephants in America, The Scientific Monthly, October
1952, 75:215-221.
4/AE Nordenskjöld. The Voyage of the Vega. London 1881.
5/Aelianus, De Natura Animalium: Kirja 2, luvut 16, 21 ja 26; 6: 4, 17, 21, 22
ja 63; 7:11; 10:25 ja 48; 11:2 ja 17; 12:39; 15:21;16:39. Aristoteles, History
of Animals, Kirja 8, luku 27; kirja 9, luku 2, jae 3; 6:20:12; 9:7:3. Parts of
Animals 4:13. Beowulf. Penguin Classics. Harmondworth, s. 112-113. Herodotos,
Historiateos 1-2. s. 58, 105-106 ja 239 WSOY 1997. Lucan, Pharsalia, Kirja 6,
677, Kirja 9, 726-732. Marcellinus Kirja 15, luku 5; 16:10. Plinius, Natural
History of Pliny, Kirja 3 luku 53; 8:9, 12, 14, 22, 28, 35, 41; 10:7. Strabo,
Kirja 6, luku 4, jae 16; 15:1:37. Ulyssis Aldrovandi, Serpentum et Draconum
Historiae. Bononiae, (1640 jKr). P. Belon du Mans, Portraits de quelques animaux,
poissons, serpents jne.(1557 jKr). Shu Hai King (Vuori ja Meri, 1122 eKr). Edes
Steven Jay Gould ei malta olla viittamatta anekdootinomaisesti pitkään ja
klassiseen kuvaukseen kielenkääntäjien painajaisista "behemotista" ja "leviatanista"
Jobin kirjan luvuissa 40-41. The Panda’s Thumb s. 259.
6/Simon Conway Morris (2000). Evolution: Bringing Molecules into the Fold. Cell
100, 1-11.
7/Cann RL, Stoneking M & Wilson AC (1987). Mitochondrial DNA and human evolution.
Nature 325, 31-36.
8/Parsons TJ ym. (1997) A high observed substitution rate in the human
mitochondrial control region. Nat. Genet. 15, 363-368.
9/Denver DR, Morris K, Lynch M, Vassilieva LL & Thomas WK (2000). High Direct
Estimate of the Mutation Rate in the Mitochondrial Genome of Caenorhabditis
elegans Science 289, 2342-2344.
10/Ingman M, Kaessmann H, Pääbö S & Gyllensten U (2000). Mitochondrial genome
variation and the origin of modern humans. Nature 408, 708-713.
11/Pääbö S. (1996). Invited Editorial, Mutational Hot Spots in the Mitochondrial
Microcosm. Am. J. Hum. Genet. 59, 493-496.
12/Adcock GJ, Dennis ES, Easteal S, Huttley GA, Jermiin LS, Peacock WJ & Thorne
A (2001). Mitochondrial DNA sequences in ancient Australians: Implications for
modern human origins. PNAS 98, 537-42.
13/Shen P ym (2000). PNAS 97, 7354-7359.
14/Teichmann, SA & Mitchison G (1999). Is There a Phylogenetic Signal in
Prokaryote Proteins? J Mol Evol 49, 98-107.
15/Doolittle WF (2000) Uprooting the tree of life. Sci. Am. 282 (2), 72-77.
16/Feng D-F, Cho G & Doolittle RF (1997). Determining divergence times with a
protein clock: Update and reevaluation. PNAS 94, 13028-13033.
17/Jain R, Rivera MC & Lake JA (1999). Horizontal gene transfer among genomes:
The complexity hypothesis. PNAS 96, 3801-306.
18/Gould SJ (1994). The evolution of life on the earth, Sci. Am. 271 (10),
62-69. Wonderful Life s. 57-60.
19/Morris SC (1993). The fossil record and the early evolution of the Metazoa.
Nature 361, 219-225.
20/Erwin DH, Valentine JW & Sepkoski JJ. A Comparative Study of Diversification
Events: The Early Paleozoic Versus the Mesozoic, Evolution 41, 1178.
21/Carroll RL (2000). Towards a new evolutionary synthesis. TREE 15, 27-32.
22/ Shu DG, Luo, HL, Morris, SC, Zhang, X-L, Hu, S-X, Chen, L, Han, J, Li, Y &
Chen, L-Z. (1999). Lower Cambrian vertebrates from south China. Nature 402,
42-46.
23/Siegfrid Scherer & Reinhard Junker (2000). Evoluutio – kriittinen analyysi.
Suomeksi toimittanut Matti Leisola, datakirjat.
24/ Research news. Dino DNA: The Hunt and the Hype. Science 261, 160-162.
25/DeSalle R, Gatesy J, Wheeler W & Grimaldi D (1992). DNA Sequences from a
Fossil Termite in Oligo-Miocene Amber and Their Phylogenetic Implications.
Science 257, 1933-1936.
26/Lindahl T (1993). Instability and decay of the primary structure of DNA.
Nature 362, 709-715.
27/Nature (1991) 352, 381.
28/Sansom IJ, Smith MP ja Smith MM (1994). Nature 368, 591.
29/Ho TY (1965). PNAS 54, 26-31.
30/Wyckoff RWG & Davidson FD. (1976) Comp. Biochem. Physiol B. 55, 95-97.
31/Science 261, 161.
32/Science (1978) 200, 1275.
33/Stankiewicz BA ym. (1997). Science 276, 1541-1543.
34/Barreto C ym. (1993) Science 262, 2020-2023.
35/J. Paleont. (1987) 61, 198-200.
36/Cano RJ & Borucki MK (1995). Revival and identification of bacterial spores
in 25- to 40-million-year-old Dominican amber. Science 268, 1060-1064.
37/Bada JL. Racemization of Amino Acids in Nature (1982). Int. Disc. Sci. Rev.
7, 30-46.
38/New Scientist 17.5.1997 s. 7.
39/Vreeland RH, Rosenzweig WD & Powers DW. (2000). Isolation of a 250
million-year-old halotolerant bacterium from a primary salt crystal. Nature 407,
897-900.
40/Vielä 1960-luvun alussa avaruuspölyä uskottiin kertyvän Maan pinnalle 5-14
miljoonaa tonnia vuodessa. (Hans Pettersson (1960) "Cosmic Spherules and
Meteoric Dust," Sci. Amer., 202:132 ). Myöhemmin arvioita on korjattu
18000-25000 tonniin vuodessa, joskin tuloksissa on hajontaa kumpaankin suuntaan.
Pölyä toki kertyy enemmän, mutta sen Maan ulkopuolisen alkuperän todistaminen on
vaikeaa. Mitä Kuuhun tulee, niin siellä ei ole eroosiota, joten pölyn pitäisi
säilyä. Kuuhun oli lähetetty ensin miehittämättömiä aluksia ja aikaisemmat
paksua pölykerrosta ennakoineet arviot olivat jo miehitetyn lennon aikana
pääasiassa väistyneet. Käsitykseen paksusta pölykerroksesta lienee vaikuttanut
myös ikuisen poleemikon Sir Fred Hoylen tieteisromaani, jossa on fiktiivinen
kuvaus kraateriin kasautuneesta pölystä.
41/ Woese CR (2002). On the evolution of cells. Proc Natl Acad Sci U S A 99,
8742-7.
Oikoluettu Helsingin yliopiston biologian opiskelijoiden ainejärjestö
Symbioosi ry:n julkaiseman Symbiontin numerosta 2/01.
Esitetyt mielipiteet eivät sellaisenaan edusta Symbioosin virallista
kantaa.

Pelasta elämä - lahjoita verta!
Safe a Life - Donate Blood!
Jos painovoima
loppuisi NYT! niin milloin se vaikuttaisi miljardin valovuoden päässä olevassa
kohteessa? Tangentin suuntaisesti.
Valovuodet eivät ole ajan yksikkö.
Valovuosi mittaa etäisyyttä, ei historiaa.
Edes valonnopeus ei mahdollisesti olekaan vakio ja saattaa rapistua asymptoottisesti arvoon 300 000 km/s.
Mutta "gravitonia" ei ole vielä löydettykään. Se on liian nopea. Ajan lyhyt hysteria.
ûMÿMû
If Gravitation would stop to exist NOW! When would it be seen in a remote star?
Straight via tangentia.
Light years are not a measure of time or history, but of distance.
Even the speed of light may not be constant, after all, and may decay asymptotically to 300 000 km/s.
In contrast to the Photon, the "Graviton" has not been even found yet. It is so fast. The short hystery of time.